大多数细菌可能存在或至少有一部分时间处于单细胞状态。在这种单细胞状态下,细菌将更容易受到一系列威胁,包括其周围环境的物理或化学性质的简单波动,更容易受到更直接的生物威胁,如抗生素、噬菌体、拮抗细菌,甚至是捕食性单细胞真核生物。因此,每一个细菌细胞都必须具备应对这种威胁的能力,微米尺度上单细胞生命的所受压力和威胁是形成细菌进化的主要力量。考察拮抗作用在自然群落中的研究表明,拮抗作用可以发挥多种作用,从促进原始生境的定殖到维持细菌群落的稳定性。
细菌与其环境相互作用的研究历史上一直集中在获取营养和抵抗非生物胁迫的策略上。这种关注淡化了细菌生命的第三个方面,而这是它们存在的同等重要的方面:即,拮抗细菌对生存的威胁。刚刚,发表在Current Biology上的一篇来自美国华盛顿大学医学院的一篇题为:“The Central Role of Interbacterial Antagonism in Bacterial Life”的文章对这一问题进行详细的讨论和论证,证明了细菌间拮抗作用的普遍性和重要性。细菌间拮抗途径的多样性和普遍性越来越明显,细菌间毒素解除其靶标的隐蔽方式强调了这些过程的高度进化性。拮抗途径的普遍性必然与同样广泛的防御策略相匹配。这些与特征明确的中央应激反应途径重叠,突出了细菌相互作用对塑造细胞生理的贡献。

背景:
一种细菌可能比另一种典型的细菌具有同等甚至更高的地位,并对这种典型的细菌来造成威胁。在一项综合分析中,考虑并权衡了地球上细菌的主要栖息地,得出的结论是,20-80%的细菌是表面相关的,并且证明了在细菌小尺度上的多样性,常常是细菌物种之间直接的细胞-细胞接触。这些观察结果延伸到栖息地,如土壤、深海和大陆亚结构,人类肠道和口腔。因此,存在一个合理的“物理证据”,表明细菌通常彼此紧邻生活。 现在,要了解在这些相互作用过程中发生了什么,即它们在本质上是拮抗的还是合作的,我们必须从体外实验,基因组证据和理论模型中进行推断。
拮抗途径的多样性和普遍性
自二十世纪初从链霉菌属中分离出首批抗生素以来,人们就已经意识到细菌杀死或抑制其他细菌生长的能力。
然而,随着细菌学逐渐发展成为一门专注于纯培养中有机体研究的学科,细菌间相互作用的表征不再受欢迎,细菌拮抗作用的研究也主要局限于寻找与临床相关的抑制分子。在21世纪早期,人们发现细菌也可以通过特殊分泌系统的作用,用抗菌毒素来对抗密切接触的邻居。很快就发现这种机制很普遍,这促使了细菌间拮抗作用研究的复兴。
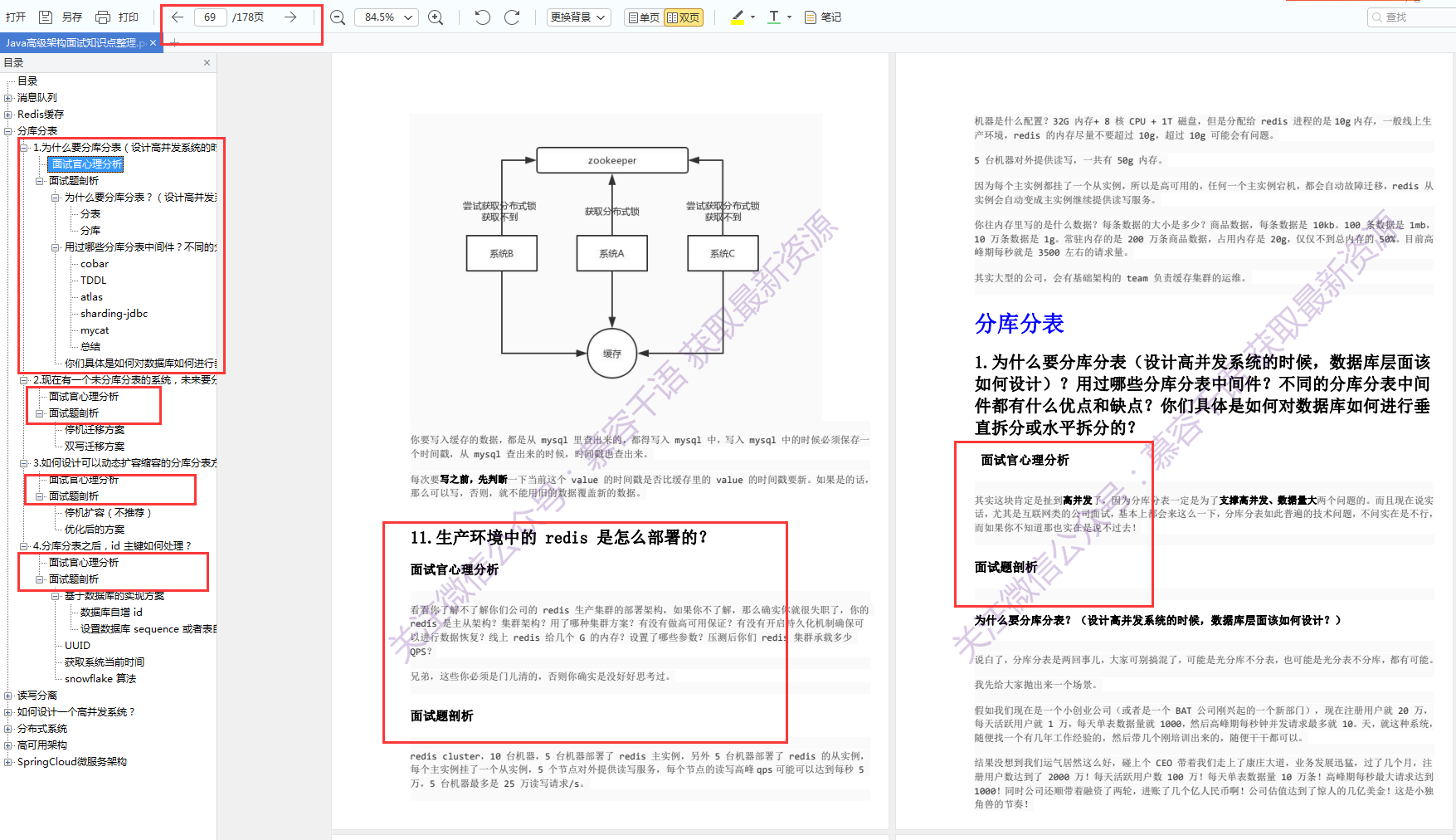
随着研究兴趣的增加和基因组数据的大量涌入,发现细菌间拮抗机制的速度正在增加。所有主要的细菌门现在已经被证明具有拮抗途径,在许多情况下包括接触依赖和非依赖机制(表1和表2)。确定的可扩散毒素包括经典的小分子抗生素,它们是链霉菌的研究重点,也包括蛋白质毒素,其大小从肽到多亚基组合。接触依赖性拮抗作用由多种特殊分泌系统介导,包括革兰氏阴性菌的IV型、V型和VI型途径以及革兰氏阳性菌的Esx分泌系统。这些系统中的每一个都向邻近细胞传递毒性效应蛋白,并利用同源免疫蛋白来防止自身和亲属中毒。
接触依赖性的细菌间拮抗作用也可以通过其他途径发生,如外膜融合介导的粘液球菌毒素交换,茎杆菌中淀粉样细菌素的表面相关产生,以及芽孢杆菌中带有羧基末端毒素结构域的肽聚糖锚定YD重复蛋白的产生。
从整个系统发生学的角度来看,不仅细菌间的拮抗机制是多种多样的; 单个物种本身可以编码多方面的拮抗武库。
多样化发生在许多层面上,包括携带多种独特拮抗机制的物种(图1),给定机制的非冗余形式(即多种毒素输出分泌途径)以及通过单个传递系统传递的过多效应子。 在生物体中发现的拮抗途径之和可占细胞总编码能力的重要部分。例如,铜绿假单胞菌编码至少六种不同的使竞争对手中毒的方法,它们共同构成了基因组的190 kb(3%)(图1A)。其中一些包含相关系统的非冗余版本,包括三个VI型分泌系统(T6SS),每个系统与多达七个独特的分泌效应子相关。 该细菌还拥有编码两种接触依赖性抑制(CDI)和三种可扩散的蛋白质毒素(称为pyocins)的基因。 革兰氏阳性枯草芽孢杆菌编码相似而广泛但不重叠的拮抗机制(图1B)。
细菌间毒素的潜伏性
细菌间拮抗途径的有效性不仅仅是其毒素内在效力的函数。毒素必须克服由细胞防御系统产生的耐受性,这些防御系统因其造成的损害而被激活,作用于细胞质分子的毒素必须突破细胞包膜,而成功的中毒策略应该相对不受抵抗力的影响。
许多抗菌毒素作用于不易从细胞外部进入的分子。 例如,似乎大多数T6SS毒素在周质中起作用,尽管有一些争论,但证据表明至少在铜绿假单胞菌中,该分泌装置最初将其抗菌蛋白传递至该隔室。 因此,具有细胞质靶标的T6SS底物需要一种穿过细菌内膜的手段。尽管这些蛋白的胞质进入细节仍然未知,但是T6SS毒素似乎并不需要通过接触依赖性抑制(CDI)系统传递毒素的特定内膜受体。
确实,铜绿假单胞菌的一种T6SS毒素Tse6似乎通过一种相当独特的机制克服了这一障碍。 该毒素可降解基本的细胞内代谢产物烟酰胺腺嘌呤二核苷酸(NAD),与细胞中最丰富的蛋白质之一,延伸因子Tu(Ef-Tu)紧密相互作用。 这种相互作用对于生化活性和毒素分泌都是必不可少的,但是破坏Tse6-Ef-Tu相互作用会使蛋白质完全无法作用于受体细胞。缺乏详细的机理理解,但简约产生了一种模型,其中Ef-Tu相互作用可能通过在细胞质内动态捕获Ef-Tu-相互作用基序来促进内膜上的Tse6易位。
通过T6SS,T4和Esx分泌系统传递毒素需要长时间的细胞间接触,这阻止了在流体条件下使用这些拮抗机制。 相反,由T5SS介导的CDI途径可以在液体培养物中生长的细胞之间起作用。 CDI毒素共有的一系列独特特征,它们在与靶细胞发生相对短暂的接触后介导其摄取。 CDI毒素以前被描述为“棒上的毒素”,因为它们由一个延伸的氨基末端细丝组成,该细丝通过b-barrel蛋白和一个羧基末端固定在分泌部位的生产细胞上 毒素结构域传递到细胞。 该报道表明,在没有靶细胞的情况下,在挤出毒素或相邻的FHA-2结构域之前,细丝的分泌会停止。 这导致发夹形结构的表面呈现,该结构由毒素的丝状部分的前半部分组成,该部分的末端具有负责靶细胞中受体结合(RBD)的域,而其余部分则由 蛋白在分泌之前向分泌通道延伸或位于周质中。 当RBD与其靶细胞上的受体结合时,分泌停滞得以缓解,并且FHA-2结构域与靶细胞的外膜缔合并嵌入其中。这随后促进了羧基末端毒素结构域向靶细胞周质的移位。 毒素中特定的脯氨酸丰富结构域被证明与分泌停滞有关。值得注意的是,缺乏该结构域的细胞在固体培养基生长条件下仍能够使靶标中毒,但丧失了在液体培养中起作用的能力。 这项研究说明了抗菌毒素访问其靶分子的途径如何对有效递送它们的条件产生深远的影响。
毒素的独创性远远超出了它们到达目标的阴险方式;如果没有办法颠覆抵抗的出现,毒素将很快被解除武装。一种“数量上的优势”方法,即通过不同机制同时传递多种毒素,可能在一定程度上已经在接触依赖型毒素传递系统中进化出来,以解决这一问题。这种策略还有一个额外的优点,即允许毒素协同作用,并便于在一系列环境条件下中毒,否则这些环境条件可能会使单一毒素的活性失效。
个别毒素也可以拥有自己的抗性颠覆解决方案。 例如,Serratia proteamaculans的ADP-核糖基酶和T6SS底物Tre1所采用的主要中毒机制是通过修饰细胞分裂蛋白FtsZ来阻止细胞分裂。然而,Tre1的活性并不局限于这个靶点;它对核糖酯表面可接近的精氨酸残基的随意修饰能力使得Tre1能够修饰其他一些必需的蛋白质。因此,如果FtsZ获得了一个突变,使其对修饰具有抵抗力,那么Tre1原则上仍然可以通过其他方式使细胞失活。
自然环境中的细菌间拮抗作用
尽管基因组证据和实验室研究对跨域细菌拮抗机制的普遍性毫无疑问,但这些机制在自然环境中的作用才刚刚开始受到关注。生物体之间的相互作用很难在原位进行监测,特别是在高度多样性的生境中或在难以实时访问的地点,如哺乳动物的肠道中。
尽管如此,采用一系列方法和不同模型系统的研究已经开始出现,这表明拮抗作用可能是细菌栖息地的一个普遍特征。
土壤包含地球上最多样化的微生物组合之一,长期以来被认为是具有拮抗能力的有机体。最近,来自特征不明显的Acidobacteria, Verrucomicrobia, Gemmatimodetes 和 Rokubacteria的土壤细菌被证明编码多种途径,以产生可作为抗菌药物的次生代谢物。然而,在土壤细菌中也经常检测到诸如交叉取食和降解协同作用等合作特性,这导致一些人质疑这种环境中拮抗作用的作用。经典生态学理论预测,两种生物所居住的生态位越重叠,它们就越有可能参与竞争行为,这就导致了这样一种假设,即土壤细菌之间的拮抗作用在邻近的相关生物之间最为强烈。有两类细菌,链霉菌属(Streptomyces sp.)和荧光假单胞菌属(Pseudomonas
fluorescens)两类细菌,它们能产生小分子抗菌剂,并能参与由细菌素介导的拮抗作用。来自同一采样点的链霉菌对比从远端采集的成对链霉菌更可能表现出拮抗作用,如果它们表现出相似的营养需求。
与链霉菌一样,具有相似营养需求的荧光假单胞菌菌株之间的相互抑制作用比那些有不同需求的菌株更强。值得注意的是,这两项研究都发现,抵抗对抗性的能力比积极抑制另一种菌株的能力更广泛。
从相关土壤细菌之间的相互作用得出了两个进化结果:获得对竞争菌株产生的毒素的抗性和/或营养专业化以避免拮抗作用。对其他细菌种群的研究将有助于解读这些细菌是否是土壤有机体中的普遍现象。
考虑到微生物在哺乳动物肠道中的定殖密度,一系列的拮抗途径是由栖息在这个生态系统中的细菌编码的就不足为奇了。然而,在没有扰动的情况下,肠道微生物群落非常稳定,这就提出了一个问题,即在这种环境中如何以及何时使用拮抗机制。这可能是肠道内的拮抗作用对于介导婴儿最初定植过程中的细菌间竞争至关重要。对拟杆菌属中T6SS基因丰度的宏基因组数据集的分析似乎支持这一点。 脆弱的芽孢杆菌特异性T6SS在婴儿定殖菌株中比在成人中更常见。使用生生物小鼠和模型社区的实验支持了该系统在肠道定植过程中的作用。的确,脆弱芽孢杆菌的T6SS有助于对生食性小鼠中敏感菌株的定植抗性。
然而,对成年肠道群落中包括T6SS在内的拮抗途径的测量表明,它们还起着最初定殖以外的作用。事实上,脆弱的芽孢杆菌的T6SS有助于gnotobiotic小鼠对敏感菌株的定殖抗性。此外,另外两种类杆菌T6SS及其各自的效应器编码在移动元件上,证据表明,在同一宿主上定殖的菌株通过水平转移获得和保持这些元件具有很强的选择性压力,从而导致效应器和免疫基因对的相容性。
在成熟的肠道菌群中,支持拮抗作用至关重要的其他证据来自获得性细菌防御(AID)和重组相关的AID(rAID)系统的发现。 这些由孤儿免疫基因组成的拟杆菌属元素普遍存在于成年肠道元基因组中,并且能够完全中和通过拮抗拟杆菌属菌株而递送的相应毒素。有趣的是,rAI簇显示出活跃基因获取的特征,表明它们可以起到适应性免疫功能的作用-与CRISPR阵列类似,但可以防御细菌间的拮抗作用而不是噬菌体攻击。
与rAID簇中编码的许多基因最接近的同源物是与T6SS以外的毒素传递途径相关的免疫基因,这些基因在拟杆菌属的外部发现,这增加了该系统对不同拮抗剂提供保护的可能性。 这些孤儿免疫基因簇在肠道细菌中的流行表明,对局部产生的毒素的抗性的获得促进了原本不相容的菌株之间的共存,这反过来又可能有助于群落的稳定。
进一步的证据支持成熟肠道菌群中拮抗作用的关键性质来自获得性细菌间防御(AID)和重组相关的AID(rAID)系统的发现。这些由孤儿免疫基因组成的类杆菌素广泛存在于成人肠道异源基因组中,能够完全中和通过拮抗拟杆菌菌株传递的相应毒素。有趣的是,rAID簇显示了活跃基因获取的特征,这表明它们可以发挥适应性免疫功能-类似于CRISPR阵列,但用于防御细菌间的对抗而不是噬菌体攻击。与rAID簇中编码的许多基因最接近的同源基因是与毒素传递途径相关的免疫基因,而T6SS则是在类杆菌之外发现的,这增加了系统对不同拮抗剂提供保护的可能性。这些孤儿免疫基因簇在肠道类杆菌中的流行表明,获得对局部产生的毒素的抵抗力促进了其他不相容菌株之间的共存,这反过来可能有助于群落的稳定。
作为可扩散抗微生物剂在成熟肠道菌群中作用的一个例子,已经确定了一个四元细菌联合体,它对小鼠的抗万古霉素肠球菌(VRE)具有定植抵抗力。这种预防作用需要革兰氏阳性菌Blautia Producta产生一种广谱的特异性抗菌肽。强调定植抗性可以在临床中发挥重要作用,接受造血细胞移植的患者对VRE感染敏感,他们拥有生产这种抗生素的基因。
革兰氏阴性菌产生的可扩散抗菌剂同样在肠道菌群中起重要作用。拟杆菌分泌抗菌蛋白,称为类杆菌分泌的抗菌蛋白(BSAP),针对靶细胞上的特定表面分子。对BSAPs的抗性是广泛存在的,并且通过与毒素基因相邻的其他靶基因的表达来介导。
在人类肠道易位基因组中,当存在BSAP基因时,具有抗性目标等位基因的菌株会富集;因此,就像由拟杆菌的T6SS介导的拮抗作用一样,这些生物体产生的可扩散的抗菌蛋白可能会在定植于不同个体的菌株之间进行兼容性选择。
上面描述的例子强调了细菌拮抗作用在原始生境定殖和捍卫已建立种群中的潜在作用。 研究表明,这些途径的第三个作用在于介导入侵。肠霍乱弧菌,霍乱弧菌和伤寒沙门氏菌均编码T6SS,有助于肠道菌落定植或表现出针对体内的共生物种。细菌素也可以介导肠道的侵袭。产质粒编码细菌素的粪肠球菌能够特异性地侵入并取代本土的粪肠球菌,并且通过改变共生微生物群落,由单核细胞增生性李斯特菌的强毒菌株产生细菌素可促进小鼠肠道定植[93]。而单核细胞增生Listeria monocytogenes菌强毒株产生的细菌素通过改变共生微生物群落来促进小鼠肠道定殖。
拮抗机制可以与外界干扰协同作用,促进病原菌入侵。发现鼠伤寒沙门氏菌的肠杆菌科特异性细菌素colicin 1b有助于其仅在炎症期间与小鼠肠道中的共生大肠杆菌株竞争,这通常会破坏肠道群落结构并促进蛋白细菌种类的生长。有趣的是,在这种情况下,肠道共生物种可以采取反策略。大肠杆菌Nissle通过产生称为微素的抗菌肽来抑制入侵的沙门氏菌的生长,这种抗菌肽是响应低铁而引起的,这是发炎的肠道的标志。此外,微素通常与铁载体偶联,从而促进铁载体被靶细胞中的铁载体吸收,而靶细胞必须主动清除铁才能维持其活性。
由于实验研究已提供证据证明细菌间的拮抗作用是微生物群落中普遍存在的一个方面,因此关于拮抗相互作用如何影响群落动态的问题也就出现了。报告表明,许多栖息地中存在的高水平细菌多样性可以通过合作代谢交叉喂养来解释。然而,这些研究往往未能明确解释活性拮抗机制,而只关注营养素的竞争。
关于合作以及拮抗和剥削性细菌间竞争对人类肠道群落动力学影响的数学模型表明,与直觉相反,拮抗作用促进稳定性。在这项分析中,合作是不稳定的,因为它有可能在物种之间建立依赖关系,从而对群落产生整体不稳定的影响。相反,对抗促进了广泛多样性水平的稳定。这项研究还确认了空间隔离是一种通过抑制物种相互作用程度来促进群落稳定的机制。
最近对鱿鱼共生菌Vibrio fischeri 的T6SS系统在寄主定殖过程中的功能的研究提供了一个真实例子,说明空间隔离和拮抗协同作用,以构建微生物群落。在这个例子中,作者发现编码T6SS的V. fischeri菌株和那些易中毒的菌株可以在同一种动物中共同定居,但只能通过占据鱿鱼发光器内不同的、空间隔离的隐窝。
对抗性防御
如果细菌间的拮抗作用像上面描述的基因组学、机械学、原位和理论研究所表明的那样普遍存在于微生物生命中,那么细菌也会进化出许多防御这些攻击的方法。一种潜在的防御机制仅仅是为了发动更好的进攻,这可能是导致对抗机制多样化和多样化的一个因素,而这些机制是由单个菌株编码的。一个有机体可以采用的相关策略是,根据特定竞争对手存在时特定活动的选择性优势,对其产生的毒素进行编码。后一种方法的一个例子是Rhs毒素。这些大的,多态性毒素由一个与羧基末端毒素相连的重复区域组成。
然而,这些对缺乏分泌所需毒素的重复部分。在鼠伤寒沙门氏菌中,这种细菌的反复传代选择了一个带有Rhs毒素的克隆,其中祖先的CT域被下游的CT域取代。这种菌株的选择是通过破坏编码这种CT毒素及其免疫决定因子的开放阅读框来实现的。尽管重组介导的毒素结构域交换尚未在该系统中得到实验验证,但编码奈瑟菌物种多态性MafB毒素的基因簇显示出类似的结构。
第二种颠覆对抗作用的方法是获得免疫决定因子,这些决定因子能够保护机体免受特定毒素的侵害(图2)。如上所述,这似乎是肠道拟杆菌的普遍特征。虽然目前为止,这些生物是唯一一个编码免疫基因阵列的群体,这些基因似乎是主动获得的,并且已经被功能性地描述过了,其他一些被称为“孤儿”免疫基因的例子也被描述过,这些基因被预测能够保护它们免受非编码它们的生物体制造的毒素的侵害。
此外,与土壤中的生产者物种相比,土壤中具有抗小分子抗生素抗性的基因更为丰富,这表明它们在保护目标细胞方面发挥着更广泛的作用。
众所周知,在临床环境中,细菌应激反应会导致抗生素耐药性。然而,这在很大程度上被认为是其提供非生物应激源保护作用的真正功能的不幸副作用。
在广泛抗菌拮抗证据的背景下重新审视这些途径,我们认为提供保护免受生物攻击实际上可能是它们在自然界的主要功能。压力反应途径的例子支持了这一点,这些途径提供了复杂的方法来对抗其他细菌产生的特定毒素的作用。例如,土壤细菌枯草芽孢杆菌(Bacillus subtilis)利用一系列互补机制来抵抗镧系抗生素(由许多革兰氏阳性细菌产生的多环肽抗菌剂)的中毒Bacillus subtilis对镧系抗生素的防御措施包括合成带负电荷的脂肪酸,它能减缓阳离子肽穿过细胞壁的速度,产生膜信号肽酶SppA,被认为有助于肽抗生素在细胞膜中降解,诱导噬菌体休克蛋白同源物,保护膜不受造孔镧系物的影响。
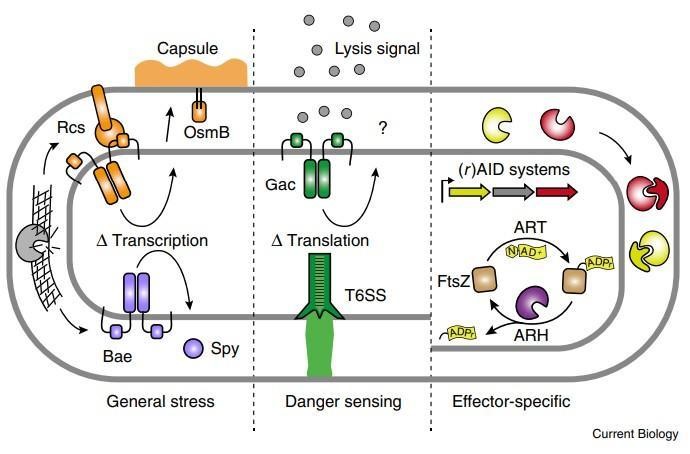
另一条将应激反应途径与细菌间拮抗联系起来的证据是来自另外一篇研究中,他们称之为竞争感应。这些作者提出了这样一个假设:细胞损伤表明存在拮抗竞争对手,诱导拮抗机制与修复系统协同提供了一种同时抵抗和反击攻击的手段。在铜绿假单胞菌中,应激反应机制和拮抗途径也受到协同调节,但由一种被称为副(绿脓杆菌对拮抗作用的反应)的不同机制。由溶解性抗菌毒素活性释放的细胞内自源性内容物作为旁分泌信号,刺激附近细胞中Gac/Rsm全球转录后调节通路的激活(图2)。随之而来的是T6SS的引入,以及共同提高对抗敌手竞争能力的其他因素。Gac / Rsm调节子中的许多基因没有已知的功能,这表明在这种生物体中尚需表征提供对抗拮抗作用的其他机制。
结论
细菌间拮抗途径的多样性和广泛分布,这些机制有助于细菌适应环境和在无数环境中竞争生存。这一点很重要,因为任何试图改造细菌群落(包括肠道微生物群落)以改变其特性的尝试,都必须包括一种方法,使引进的生物体能够与它们总是会遇到的其他菌相互作用进行抗衡。拮抗途径多样化的一个更微妙的潜在结果是,它们有可能成为毒素的进化库,包括细菌病原体用来对抗哺乳动物宿主的毒素,以及真核生物水平获得的抵御细菌的毒素。
表1 细菌拮抗作用的非接触依赖性机制。
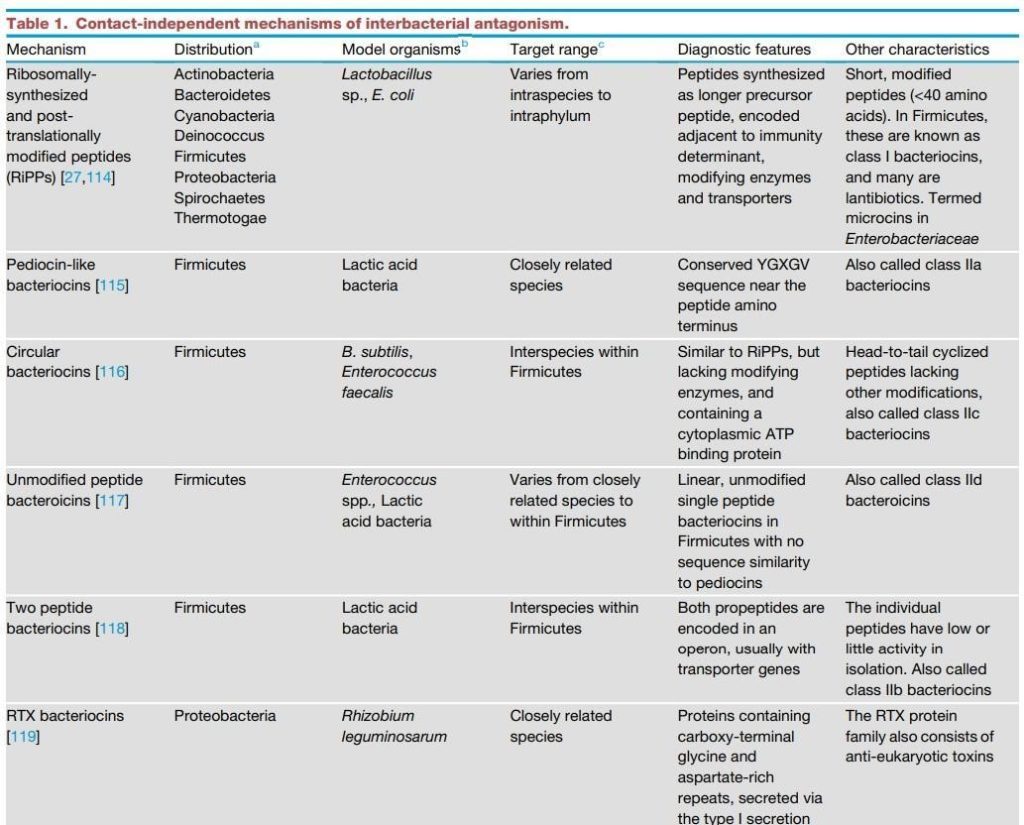
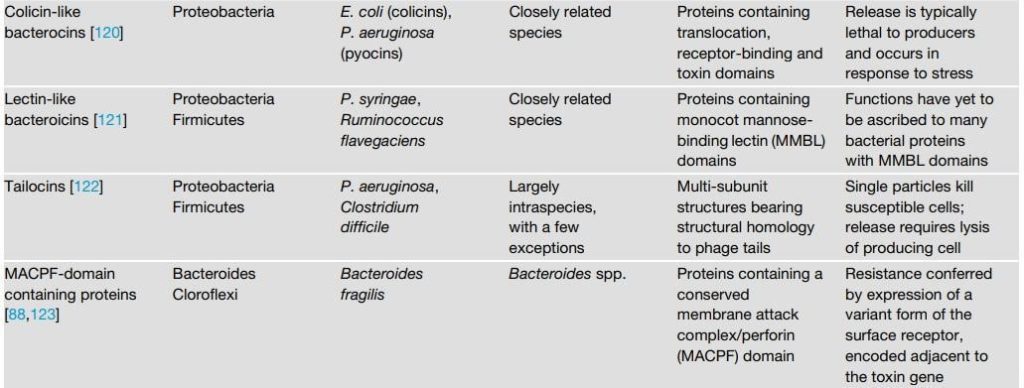

表2 接触依赖性细菌拮抗途径
图1 细菌基因组中充满了编码各种细菌间拮抗途径的基因座。
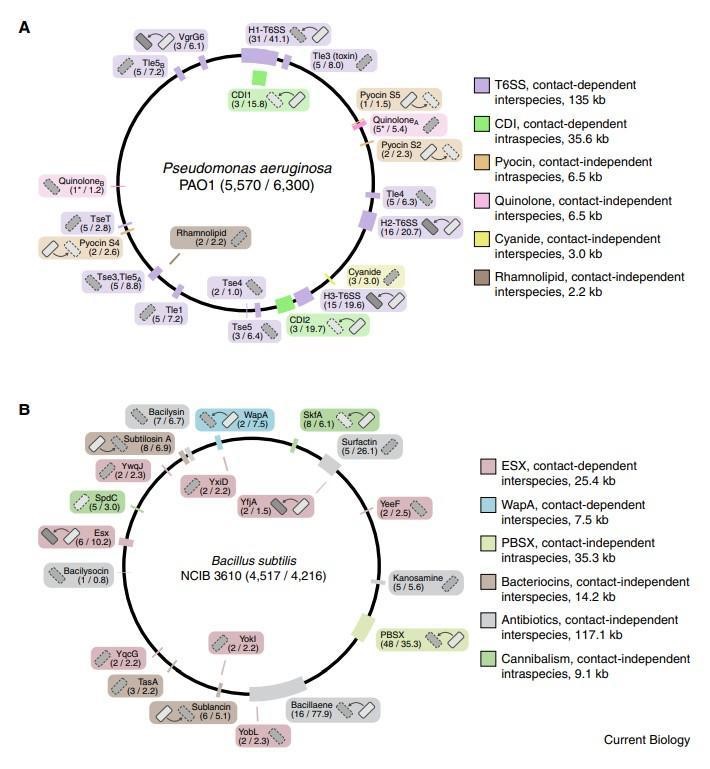
图2 革兰氏阴性细菌对T6SS的多种防御途径。
本文转载自谷禾健康